Rregullimi i sintezës së kolesterolit
Rregullimi i sintezës së kolesterolit - enzima e tij kryesore (reduktaza HMG-CoA) kryhet në mënyra të ndryshme.
Fosforilimi / deposforilimi i reduktazës HMG. Me një rritje të raportit të insulinës / glukagonit, kjo enzimë fosforilon dhe kalon në një gjendje aktive. Veprimi i insulinës kryhet përmes 2 enzimave.
HMG-CoA reduktaza kinazë fosfatazë, e cila e shndërron kinazën në një gjendje deposferizuese joaktive:
Reduktaza e fosfotazës HMG-CoA duke e shndërruar atë në një gjendje aktive të deposferuar. Rezultati i këtyre reaksioneve është formimi i një forme aktive të deposferuar të reduktazës HMG-CoA.
Si pasojë, gjatë periudhës së përthithjes, kolesteroli rritet. Gjatë kësaj periudhe, disponueshmëria e substratit fillestar për sintezën e kolesterolit - acetil - CoA rritet (si rezultat i ngrënies së ushqimit që përmban karbohidrate dhe yndyrna, pasi që acetil CoA formohet gjatë prishjes së glukozës dhe acideve yndyrore).
Në gjendjen pasabsorbente, glukagoni përmes proteinogjenazës A stimulon fosforilimin e HMG - CoA - reduktazës, duke e shndërruar atë në një gjendje joaktive. Ky veprim përmirësohet me faktin se në të njëjtën kohë glukagoni stimulon fosforilimin dhe inaktivizimin e fosfotazës së reduktazës HMG-CoA, duke e mbajtur kështu reduktazën HMG-CoA në një gjendje joaktive fosforiluese. Si rezultat, sinteza e kolesterolit në periudhën pasabsorbimit dhe gjatë agjërimit frenohet nga sinteza endogjene. Nëse përmbajtja e kolesterolit në ushqim u soll në 2%, atëherë sinteza e kolesterolit endogjen u ul ndjeshëm. Por ndërprerja e plotë e sintezës së kolesterolit nuk ndodh.
Shkalla e frenimit të biosintezës së kolesterolit nën ndikimin e kolesterolit që vjen nga ushqimi ndryshon nga personi në person. Kjo tregon individualitetin e proceseve të formimit të kolesterolit. Duke zvogëluar intensitetin e sintezës së kolesterolit, është e mundur të zvogëlohet përqendrimi i tij në gjak.
Nëse ekuilibri midis konsumit të kolesterolit me ushqim dhe sintezës së tij në trup nga njëra anë dhe sekretimit të acideve biliare dhe kolesterolit nga ana tjetër është i prishur, përqendrimi i kolesterolit në indet dhe gjaku ndryshon. Pasojat më të rënda shoqërohen me një rritje të përqendrimit të kolesterolit në gjak (hiperkolesterolemia), ndërsa gjasat e zhvillimit të aterosklerozës dhe kolelithiasis rritet.
Hiperkolesterolemia familjare (HCS) - kjo formë është shumë më e zakonshme - rreth 1 pacient për 200 persona. Një defekt i trashëguar në HCS është një shkelje e thithjes së LDL nga qelizat, dhe për këtë arsye, një rënie në shkallën e katabolizmit LDL. Si rezultat, përqendrimi i LDL në gjak rritet, si dhe kolesterolit pasi ka shumë në LDL. Prandaj, me HCS, depozitimi i kolesterolit në inde, në veçanti në lëkurë (xanthoma), në muret e arterieve është karakteristik.
Pengimi i sintezës së reduktazës HMG-CoA
Produkti përfundimtar i rrugës metabolike të kolesterolit. Ajo zvogëlon shkallën e transkriptimit të gjenit HMG-CoA reductase, duke penguar kështu sintezën e vet. Mëlçia po sintetizon në mënyrë aktive acidet biliare nga kolesteroli, dhe për këtë arsye acidet biliare frenojnë aktivitetin e gjenit HMG-CoA reductase. Meqenëse reduktaza HMG-CoA ekziston pas sintezës së rreth 3, frenimi i sintezës së këtij enzima të kolesterolit është një rregullim efektiv.
Nuk e gjetët atë që ju po kërkoni? Përdorni kërkimin:
Shkëmbimi i kolesterolit ester
Fondi i kolesterolit përmban estere të kolesterolit dhe kolesterolit, të cilat gjenden si në qeliza ashtu edhe në lipoproteinat në gjak.
Pjesa II Metabolizmi dhe energjia
Në qelizat, esterifikimi i kolesterolit ndodh me veprimin e acil-CoA-kolesterol-aciltransferazë (AChAT):
Acil-CoA + Kolesterol - * HS-KoA + Acilkolesterol
Në qelizat njerëzore, linoleylcholesterol është formuar kryesisht. Për dallim nga kolesteroli i lirë, esterët e tij në membranat qelizore përmbajnë në sasi shumë të vogla dhe gjenden kryesisht në citosol si pjesë e pikave të lipideve. Formimi i esterëve mund të konsiderohet, nga njëra anë, si një mekanizëm për heqjen e kolesterolit të tepërt nga membranat, dhe nga ana tjetër, si një mekanizëm për ruajtjen e kolesterolit në qelizë. Mobilizimi i burimeve ndodh me pjesëmarrjen e enzimave të esterazës që hidrolizojnë eseret e kolesterolit:
Acilkolesterol + H 2O - * Acid yndyror + Kolesterol
Sinteza dhe hidroliza e estereve ndodh në shumë qeliza, por është veçanërisht aktive në qelizat e korteksit veshkor: në këto qeliza deri në 80% të të gjithë kolesterolit përfaqësohet nga estrët, ndërsa në qelizat e tjera zakonisht është më pak se 20%.
Formimi i esterëve në liponroteinat në gjak bëhet me pjesëmarrjen e aciltransferazës së lecithin-kolesterolit (LHAT), e cila katalizon transferimin e mbetjeve acil nga pozicioni i i leqitinës në kolesterol (Fig. 10.31) LHAT formohet në mëlçi, sekretohet në gjak dhe bashkangjitet lipoproteinave. Shkalla e esterifikimit për lipoproteinat e ndryshme është dukshëm e ndryshme dhe varet nga prania e apolipoproteinave që aktivizojnë LHAT (kryesisht apo-AT, si dhe CI) ose frenojnë (C-II) këtë enzimë.LHAT më aktive në HDL, në të cilën apo-AT duhet te olee 2/3 e gjitha proteinat. Nderon më të mëdha coli esters e acideve oleik dhe linoleic formuar. Të lipoproteina tjera ester formacioni ndodh në një shkallë të ngadaltë se në HDL.
Fig. 10.31. Formimi i estrave të kolesterolit nën veprimin e LHAT
LHAT lokalizohet në shtresën sipërfaqësore të HDL dhe përdor kolesterolin në monolayer fosfolipid si një substrat. Esteret e kolesterolit të formuar këtu, për shkak të hidrofobicitetit të tyre të plotë, mbahen dobët brenda
Kapitulli 10. Metabolizmi dhe funksionimi i lipideve
monolayer fosfolipide dhe zhytur në thelbin lipidik të lipoproteinës. Në të njëjtën kohë, një vend për kolesterol çlirohet në monolerin fosfolipid, i cili mund të mbushet me kolesterol nga membranat qelizore ose nga lipoproteinat e tjera. Kështu, HDL duket se është një kurth kolesterol si rezultat i veprimit të LHAT.
Sinteza e acideve biliare
Në mëlçi, një pjesë e kolesterolit shndërrohet në acide biliare. Acidet galike mund të konsiderohen si derivate të acidit kolanik (Fig. 10.32).
Acidi kolanik si i tillë nuk është formuar në trup. Në hepatocitet, kolesteroli drejtpërdrejt prodhon acide kenodeoksikolike dhe kolike - acide biliare parësore (Fig. 10.33, shiko gjithashtu Fig. 10.12).
Biosinteza e kolesterolit
Biosinteza e kolesterolit ndodh në retikulumin endoplazmatik. Burimi i të gjitha atomeve të karbonit në molekulë është acetil-SCoA, i cili vjen këtu nga mitokondria si pjesë e citrateve, si në sintezën e acideve yndyrore. Biosinteza e kolesterolit konsumon 18 molekula ATP dhe 13 molekula NADPH.
Formimi i kolesterolit ndodh në më shumë se 30 reaksione, të cilat mund të grupohen në disa faza.
1. Sinteza e acidit mevalonik.
Dy reagimet e para të sintezës përkojnë me reaksionet e ketogjenezës, por pas sintezës së 3-hidroksi-3-metilglutaryl-ScoA, enzima hyn reduktaza hidroksimetil-glutaryl-ScoA (Reduktaza HMG-SCOA), duke formuar acidin mevalonik.
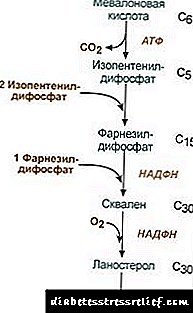 |
Skema e reaksionit të sintezës së kolesterolit2. Sinteza e difosfatit izopentenil. Në këtë fazë, tre mbetje fosfat janë ngjitur në acidin mevalonik, atëherë është dekarboksiluar dhe dehidrogjenuar. 3. Pas kombinimit të tre molekulave të isopentenil difosfatit, sintetizohet farnesil difosfati. 4. Sinteza e squalenit ndodh kur lidhen dy mbetje farnesil difosfat. 5. Pas reaksioneve komplekse, skaleni linear ciklizon në lanosterol. 6. Largimi i grupeve të tepërta metilike, restaurimi dhe izomerizimi i molekulës çon në shfaqjen e kolesterolit. Rregullimi i veprimtarisë së reduktazës hidroksimetilglutaryl-S-CoA3. Shkalla e biosintezës së kolesterolit varet gjithashtu nga përqendrimi proteina specifike bartëseduke siguruar lidhjen dhe transportimin e metabolitëve të sintezës interfektive hidrofobike. Ju mund të kërkoni ose lini mendimin tuaj. Pika kryesore e rregullimit është reagimi i formimit të acidit mevalonik. 1. Rregullimi allosterik. Kolesterolit, dhe në mëlçi - dhe acidet biliare pengojnë reduktazën HMG-CoA. 2. Represioni i sintezës së kolesterolit reduktazë HMG-CoA. 3. Rregullimi me anë të fosforilimit-deposforilimit të reduktazës HMG-CoA, formë aktive jo fosforiluese. Glukagoni shkakton çaktivizim, dhe insulina shkakton aktivizim përmes një kaskade komplekse të reaksioneve. Kështu, shkalla e sintezës së kolesterolit ndryshon me një ndryshim në gjendjet absorbuese dhe postabsorbuese. 4. Shkalla e sintezës së reduktazës HMG-CoA në mëlçi i nënshtrohet luhatjeve të qarta ditore: maksimumi në mesnatë, dhe minimumi në mëngjes. Shkëmbimi i kolesterolit ester Në qelizat esterifikimi i kolesterolit ndodh kur ekspozohet acyl-CoA kolesterol aciltransferazë (ACAT) Acil-CoA + Kolesterol ® NS-CoA + Acilkolesterol Në qelizat, formohet kryesisht linoleylkolesteroli. Estetët gjenden kryesisht në citosol si pjesë e pikave të lipideve. Formimi i esterëve mund të konsiderohet, nga njëra anë, si një mekanizëm për heqjen e kolesterolit të tepërt nga membranat, dhe nga ana tjetër, si një mekanizëm për ruajtjen e kolesterolit në qelizë. Mobilizimi i rezervave ndodh me pjesëmarrjen e enzimave esterasehidrolizues të estrave të kolesterolit: Acilkolesterol + H2Acid yndyror O Kolesterol Sinteza dhe hidroliza e estereve është veçanërisht aktive në qelizat e korteksit adrenal. Në lipoproteinat në gjak formimi ester ndodh me pjesëmarrjen e lecithin-kolesterol aciltransferazë (LHAT), duke katalizuar transferimin e mbetjeve acil nga lecithin në kolesterol. LHAT formohet në mëlçi, sekretohet në rrjedhën e gjakut dhe bashkangjitet lipoproteinave. LHAT më aktiv në HDL, ku lokalizohet në shtresën sipërfaqësore. Esteret e kolesterolit të formuar këtu janë hidrofobë dhe zhytur në thelbin lipid. Në monolayer fosfolipid, ekziston hapësirë e lirë për kolesterolin, i cili mund të mbushet me kolesterol nga membranat qelizore ose nga lipoproteinat e tjera. Kështu, HDL duket se është një kurth kolesterol si rezultat i veprimit të LHAT. Sinteza e acideve biliare Në mëlçi, një pjesë e kolesterolit shndërrohet në acide biliare. Acidet biliare mund të konsiderohen si derivate të acidit kolanik. Acidi kolanik si i tillë nuk është formuar në trup. Në hepatocitet nga kolesteroli, acidet biliare primare formohen - chenodeoxycholic dhe cholic. Pas sekretimit të biliare në zorrë nën veprimin e enzimave të florës së zorrëve prej tyre formohen acidet biliare dytësore - lithocholic dhe deoxycholic. Ato përthithen nga zorrët, me gjakun e venës së portalit hyjnë në mëlçi, dhe më pas në biliare. Bili përmban kryesisht acide biliare të konjuguara, d.m.th, përbërjet e tyre me glicinë ose taurinë. Përqendrimi i acideve biliare në biliare është rreth 1%. Pjesa kryesore e acideve biliare është e përfshirë në qarkullimi hepatoenterik.Një pjesë e vogël e acideve biliare - rreth 0,5 g në ditë - ekskretohet në feces. Kjo kompensohet nga sinteza e acideve biliare të reja në mëlçi, fondi i acideve biliare azhurnohet në rreth 10 ditë. Kolesteroli gjithashtu ekskretohet kryesisht përmes zorrëve. Ajo hyn në zorrët me ushqim dhe nga mëlçia si pjesë e biliare. Kolesteroli i përthithur në gjak përmban një fraksion me origjinë nga biliare (kolesterol endogjensintetizohet në mëlçi), dhe fraksioni që rrjedh nga ushqimi (kolesterol ekzogjen). Largimi i kolesterolit nga indet ndodh nga oksidimi i tij drejt acideve biliare në mëlçi, pasuar nga sekretimi i tyre me feces (afërsisht 0,5 g në ditë) dhe nga sekretimi i kolesterolit të pandryshuar (gjithashtu me feces). Në gjendje të palëvizshme: (Kolesterolifund + Kolesterolind) - (Kolesterolekskr + Acidet biliareekskr) = 0 Nëse ky ekuilibër është i shqetësuar, përqendrimi i kolesterolit në inde dhe në gjak ndryshon. Kolesterol i rritur në gjak - hiperkolesterolemia. Kjo rrit mundësinë e arteriosklerozës dhe sëmundjes së tëmthit. RREGULLIMI I SHKENCS S L LIPIDIT Metabolizmi i lipideve rregullohet nga sistemi nervor qendror. Që zgjat shumë stresi negativ emocional, një rritje në lëshimin e katekolaminave në qarkullimin e gjakut mund të shkaktojë humbje të dukshme të peshës. efekt glucagon në sistemin lipolitik është i ngjashëm me veprimin e katekolaminave. adrenalin dhe norepinephrine rrisin aktivitetin e lipazës së indeve dhe shkallën e lipolizës në indin dhjamor, si rezultat, rritet përmbajtja e acideve yndyrore në plazmën e gjakut. insulinë ka efektin e kundërt të adrenalinës dhe glukagonit në lipolizën dhe mobilizimin e acideve yndyrore. Hormoni i rritjes stimulon lipolizën, duke indikuar sintezën e ciklazës acenilate. Hipofunksioni i hipofizës çon në depozitimin e yndyrës në trup (mbipesha e hipofizës). Tiroksina, hormonet seksualendikojnë edhe në metabolizmin e lipideve. Largimi i gjëndrave seksuale tek kafshët shkakton depozitim të tepërt të yndyrës. SHPORDRNDARJES METABOLIKE LIPID Kolesteroli është një steroide specifike për organizmat e kafshëve. Vendi kryesor i formimit të tij në trupin e njeriut është mëlçia, ku sintetizohet 50% e kolesterolit, 15-20% formohet në zorrën e vogël, pjesa tjetër sintetizohet në lëkurë, lëvore veshkore dhe gonada. Burimet e formimit të fondit të kolesterolit dhe mënyrat e shpenzimit të tij janë paraqitur në Figurën 22.1. Fig. 22.1. Formimi dhe shpërndarja e kolesterolit në trup. Kolesteroli i trupit të njeriut (sasia totale prej rreth 140 g) me kusht mund të ndahet në tre pishina: 30 g), në shkëmbim të shpejtë, përbëhet nga kolesterol i murit të zorrëve, plazmës së gjakut, mëlçisë dhe organeve të tjera parenkimale, rinovimi ndodh në 30 ditë (1 g / ditë), 50 g), duke shkëmbyer ngadalë kolesterolin e organeve dhe indeve të tjera, 60 g), një kolesterol shumë i shkëmbyer ngadalë i palcës kurrizore dhe trurit, ind lidhës, shkalla e azhurnimit llogaritet me kalimin e viteve. Sinteza e kolesterolit ndodh në citosolin e qelizave. Kjo është një nga rrugët më të gjata metabolike në trupin e njeriut. Ajo vazhdon në 3 faza: e para përfundon me formimin e acidit mevalonik, e dyta me formimin e squalenit (struktura lineare hidrokarbure që përbëhet nga 30 atome karboni). Gjatë fazës së tretë, skaleni shndërrohet në një molekulë lanosteroli, atëherë ekzistojnë 20 reagime të njëpasnjëshme që shndërrojnë lanosterolin në kolesterol. Në disa inde, grupi hidroksil i kolesterolit esterifikon për të formuar eserë. Reaksioni katalizohet nga enzima ndërqelizore AHAT (acylCoA: aciltransferaza e kolesterolit). Reagimi i esterifikimit gjithashtu ndodh në gjak në HDL, ku ndodhet enzima LHAT (leçitina: aciltransferaza e kolesterolit). Esteret e kolesterolit janë forma në të cilën transportohet me anë të gjakut ose depozitohet në qeliza. Në gjak, rreth 75% e kolesterolit është në formën e estrave. Sinteza e kolesterolit rregullohet duke ndikuar në aktivitetin dhe sasinë e enzimës kryesore të procesit - reduktaza 3-hidroksi-3-metilglutaryl-CoA (reduktaza HMG-CoA). Kjo arrihet në dy mënyra: 1. Fosforilimi / deposforilimi i reduktazës HMG-CoA. Insulina stimulon deposforyilimin e reduktazës HMG-CoA, duke e përkthyer kështu në një gjendje aktive. Prandaj, në periudhën e përthithjes rritet sinteza e kolesterolit. Gjatë kësaj periudhe, disponueshmëria e substratit fillestar për sintezën, acetil-CoA, gjithashtu rritet. Glukagoni ka efektin e kundërt: përmes proteinës kinazë A, ajo stimulon fosforilimin e reduktazës HMG-CoA, duke e kthyer atë në një gjendje joaktive. Si rezultat, sinteza e kolesterolit në periudhën pasabsorbimit dhe gjatë agjërimit frenohet. 2. Frenimi i sintezës së reduktazës HMG-CoA.Kolesteroli (produkti përfundimtar i rrugës metabolike) zvogëlon shkallën e transkriptimit të gjenit HMG-CoA reductase, duke penguar kështu sintezën e tij, dhe acidet biliare gjithashtu shkaktojnë një efekt të ngjashëm. Transporti i kolesterolit në gjak kryhet si pjesë e ilaçit. LP sigurojnë kolesterol ekzogjen në inde, përcaktojnë rrjedhën e tij midis organeve dhe sekretimin nga trupi. Kolesterolit ekzogjen i dorëzohet mëlçisë si pjesë e mbetjes ChM. Atje, së bashku me kolesterolin endogjen të sintetizuar, formon një fond të përbashkët. Në hepatocitet, TAG dhe kolesteroli paketohen në VLDL, dhe në këtë formë sekretohen në gjak. Nën ndikimin e LP-lipazës, duke hidrolizuar TAG ndaj glicerinës dhe acideve yndyrore në gjak, VLDLPs së pari shndërrohen në STLP dhe më pas në LDLP që përmbajnë deri në 55% të kolesterolit dhe esterët e tij. LDL është forma kryesore e transportit të kolesterolit në të cilën ai dorëzohet në inde (70% e kolesterolit dhe esterët e tij në gjak janë pjesë e LDL). LDL nga gjaku hyn në mëlçi (deri në 75%) dhe inde të tjera që kanë receptorë LDL në sipërfaqen e tyre. Nëse sasia e kolesterolit që hyn në qelizë tejkalon nevojën e saj, atëherë shtypet sinteza e receptorëve LDL, gjë që zvogëlon rrjedhën e kolesterolit nga gjaku. Me një ulje të përqendrimit të kolesterolit të lirë në qelizë, përkundrazi, aktivizohet sinteza e receptorit. Hormonet janë të përfshirë në rregullimin e sintezës së receptorit LDL: insulina, triiodothyronine dhe hormonet seksuale rrisin formimin e receptorëve, dhe glukokortikoidet ulen. Në të ashtuquajturin "transporti i kthimit të kolesterolit", d.m.th. shtegu që siguron kthimin e kolesterolit në mëlçi, HDL luan rolin kryesor. Ato sintetizohen në mëlçi në formën e pararendësve të papjekur që praktikisht nuk përmbajnë kolesterol dhe TAG. Pararendësit HDL në gjak janë të ngopur me kolesterol, duke marrë atë nga LP-të e tjera dhe membranat qelizore. Transferimi i kolesterolit në HDL përfshin enzimën LHAT të vendosur në sipërfaqen e tyre. Kjo enzimë bashkon një mbetje të acideve yndyrore nga fosfatidilkolina (lecitina) te kolesteroli. Si rezultat, formohet një molekulë hidrofobe e esterit të kolesterolit, i cili lëviz brenda HDL. Kështu që, jo i dehur HDL, i pasuruar me kolesterol, shndërrohet në HDL 3 - grimca të pjekura dhe më të mëdha. HDL 3 shkëmbejnë esteret e kolesterolit për TAG të përfshira në VLDL dhe STD me pjesëmarrjen e një proteine specifike që transferon esterët e kolesterolit midis lipoproteinave. Në këtë rast, HDL 3 shndërrohen në HDL2, madhësia e së cilës rritet për shkak të akumulimit të TAG. VLDL dhe STDL nën ndikimin e LP-lipazës shndërrohen në LDL, e cila kryesisht dërgon kolesterol në mëlçi. Një pjesë e vogël e kolesterolit dorëzohet në mëlçi të HDL2 dhe HDL. Sinteza e acideve biliare. Në mëlçi, 500-700 mg acide biliare në ditë sintetizohen nga kolesteroli. Formimi i tyre përfshin reagime të futjes së grupeve hidroksil me pjesëmarrjen e hidroksilazave dhe reagimin e oksidimit të pjesshëm të zinxhirit anësor të kolesterolit (Fig. 22.2): Fig. 22.2. Skema e formimit të acideve biliare. Reagimi i parë i sintezës - formimi i 7-a-hidroksikolesterol - është rregullues. Aktiviteti i enzimës që katalizon këtë reagim pengohet nga produkti përfundimtar i rrugës, acidet biliare. Një mekanizëm tjetër rregullues është fosforilimi / deposforilimi i enzimës (forma e fosforiluar e 7-a-hidroksilazës është aktive). Rregullimi është gjithashtu i mundur duke ndryshuar sasinë e enzimës: kolesterolit shkakton transkriptimin e gjenit 7-a-hidroksilazë, dhe acidet biliare shtypin. Hormonet tiroide shkaktojnë sintezën e 7-a-hidroksilazës, dhe shtypjen e estrogjenit. Një efekt i tillë i estrogjenit në sintezën e acideve biliare shpjegon pse sëmundja e tëmthit shfaqet tek gratë 3-4 herë më shpesh sesa tek burrat. Acidet folike dhe kenodeoksikolike të formuara nga kolesteroli quhen "acide biliare primare". Pjesa më e madhe e këtyre acideve pëson konjugim - shtimi i molekulave të glicinës ose taurinës në grupin karboksil të acidit biliare. Konjugimi fillon me formimin e formës aktive të acideve biliare - derivatet e CoA, atëherë bashkohen taurina ose glicina, dhe si rezultat formohen 4 variante të konjugateve: taurokolike dhe taurohenodeoksikolike, glukokolike dhe acide glikohenodeoksikolike. Ata janë emulsifikues dukshëm më të fortë se acidet biliare origjinale. Konjugat me glicinë formohen 3 herë më shumë sesa me taurinën, pasi që sasia e taurinës në trup është e kufizuar. Në zorrë, një sasi e vogël e konjugateve të acideve biliare primare nën veprimin e enzimave bakteriale shndërrohen në acide biliare sekondare. Acidi deoksikolik, i formuar nga koliku, dhe litokoli, i formuar nga deoksikolik, janë më pak të tretshëm dhe thithin më ngadalë në zorrët. Rreth 95% të acideve biliare që hyjnë në zorrë kthehen në mëlçi përmes venës portal, më pas sekretohen përsëri në biliare dhe ripërdoren në emulsifikimin e yndyrnave. Kjo rrugë e acideve biliare quhet qarkullim enterohepatik. Me feces, acidet biliare dytësore kryesisht hiqen. Sëmundja e gurëve të tëmthit (kolelithiaza) është një proces patologjik, në të cilin formohen gurë në fshikëz e tëmthit, baza e së cilës është kolesterol. Lëshimi i kolesterolit në biliare duhet të shoqërohet me një lëshim proporcional të acideve biliare dhe fosfolipideve që mbajnë molekulat e kolesterolit hidrofobik në gjendjen mikellare. Arsyet që çojnë në një ndryshim në raportin e acideve biliare dhe kolesterolit në biliare janë: ushqimi i pasur me kolesterol, ushqimi me kalori të lartë, ngecja e biliare në fshikëz e tëmthit, qarkullimi i dëmtuar i enterohepatës, sinteza e dëmtuar e acideve biliare, infeksioni i fshikëzës së tëmthit. Në shumicën e pacientëve me kolelithiasis, rritet sinteza e kolesterolit, dhe sinteza e acideve biliare nga ajo është ngadalësuar, gjë që çon në një shpërpjesëtim të sasisë së kolesterolit dhe acideve biliare të sekretuara në biliare. Si rezultat, kolesteroli fillon të precipitojë në fshikëz e tëmthit, duke formuar një sediment viskoz që ngurtësohet gradualisht. Ndonjëherë është e mbarsur me bilirubin, proteina dhe kripëra të kalciumit. Gurët mund të përbëhen vetëm nga kolesterol (gurë kolesterol) ose një përzierje e kolesterolit, bilirubinës, proteinave dhe kalciumit. Gurët e kolesterolit janë zakonisht të bardhë, dhe gurët e përzier janë kafe në hije të ndryshme. Në fazën fillestare të formimit të gurit, acidi chenodeoksikolik mund të përdoret si ilaç. Pasi të jetë në fshikëz e tëmthit, ajo gradualisht shpërndan gurët e kolesterolit, por ky është një proces i ngadaltë që zgjat disa muaj. Atherosclerosis është një patologji që karakterizohet nga shfaqja e pllakave aterogjene në sipërfaqen e brendshme të murit vaskular. Një nga arsyet kryesore për zhvillimin e një patologjie të tillë është një shkelje e ekuilibrit midis marrjes së kolesterolit nga ushqimi, sintezës së tij dhe sekretimit nga trupi. Pacientët me arteriosklerozë kanë ngritje të përqendrimeve të LDL dhe VLDL. Ekziston një lidhje e kundërt midis përqendrimit të HDL dhe mundësisë së zhvillimit të arteriosklerozës. Kjo është në përputhje me konceptin e funksionimit të LDL si bartës të kolesterolit në inde, dhe HDL nga indet. "Parakushti" themelor metabolik për zhvillimin e arteriosklerozës është hiperkolesterolemia. (kolesterol i lartë në gjak). Hiperkolesterolemia zhvillohet: 1. për shkak të marrjes së tepërt të kolesterolit, karbohidrateve dhe yndyrnave, 2. një predispozicion gjenetik që konsiston në defekte trashëgimore në strukturën e receptorëve LDL ose apoB-100, si dhe në rritjen e sintezës ose sekretimit të apo AP-100 (në rastin e hiperlipidemisë familjare të kombinuar, në të cilën ngriten përqendrimet e gjakut dhe kolesterolin dhe TAG). Një rol të rëndësishëm në mekanizmat e zhvillimit të arteriosklerozës luhet nga modifikimi i ilaçit. Ndryshimet në strukturën normale të lipideve dhe proteinave në LDL i bëjnë ato të huaja për trupin dhe për këtë arsye më të arritshme për kapje nga fagocitet. Modifikimi i drogës mund të ndodhë nga disa mekanizma: 1. glikozilimi i proteinave që ndodh kur rritet përqendrimi i glukozës në gjak, 2. modifikimi i peroksidit, duke çuar në ndryshime në lipide në lipoproteina dhe strukturën e apoB-100, 3. formimi i komplekseve autoimune të antitrupave LP (ilaçet e ndryshuara mund të shkaktojnë formimin e autoantitrupave). LDL i modifikuar thithet nga makrofagët. Ky proces nuk rregullohet nga sasia e kolesterolit të përthithur, si në rastin e hyrjes së tij në qeliza përmes receptorëve specifikë, prandaj makrofagët mbingarkohen me kolesterol dhe shndërrohen në “qeliza shkumë” që depërtojnë në hapësirën subendoteliale. Kjo çon në formimin e njollave lipide ose shiritave në murin e enëve të gjakut. Në këtë fazë, endoteli vaskular mund të ruajë strukturën e tij. Me një rritje të numrit të qelizave të shkumës, ndodh dëmtimi endotelial. Dëmtimi kontribuon në aktivizimin e trombociteve. Si rezultat, ata sekretojnë tromboxane, e cila stimulon grumbullimin e trombociteve, dhe gjithashtu fillojnë të prodhojnë një faktor të rritjes së trombociteve, e cila stimulon përhapjen e qelizave të muskujve të lëmuar. Këto të fundit migrojnë nga mediale në shtresën e brendshme të murit arterial, duke kontribuar kështu në rritjen e pllakës. Pastaj pllaka mbin me ind fibroze, qelizat nën membranën fibroze janë nekrotike, dhe kolesteroli depozitohet në hapësirën ndërqelizore. Në fazat e fundit të zhvillimit, pllaka është e mbarsur me kripëra të kalciumit dhe bëhet shumë e dendur. Në fushën e pllakës, mpiksjet e gjakut shpesh formohen, duke bllokuar lumenin e anijes, e cila çon në shqetësim akut të qarkullimit të gjakut në vendin përkatës të indeve dhe zhvillimin e një sulmi në zemër. Rregullimi i sintezës së kolesterolit - enzima e tij kryesore (reduktaza HMG-CoA) kryhet në mënyra të ndryshme. Fosforilimi / deposforilimi i reduktazës HMG. Me një rritje të raportit të insulinës / glukagonit, kjo enzimë fosforilon dhe kalon në një gjendje aktive. Veprimi i insulinës kryhet përmes 2 enzimave. HMG-CoA reduktaza kinazë fosfatazë, e cila e shndërron kinazën në një gjendje deposferizuese joaktive: Reduktaza e fosfotazës HMG-CoA duke e shndërruar atë në një gjendje aktive të deposferuar. Rezultati i këtyre reaksioneve është formimi i një forme aktive të deposferuar të reduktazës HMG-CoA. Si pasojë, gjatë periudhës së përthithjes, kolesteroli rritet. Gjatë kësaj periudhe, disponueshmëria e substratit fillestar për sintezën e kolesterolit - acetil - CoA rritet (si rezultat i ngrënies së ushqimit që përmban karbohidrate dhe yndyrna, pasi që acetil CoA formohet gjatë prishjes së glukozës dhe acideve yndyrore). Në gjendjen pasabsorbente, glukagoni përmes proteinogjenazës A stimulon fosforilimin e HMG - CoA - reduktazës, duke e shndërruar atë në një gjendje joaktive. Ky veprim përmirësohet me faktin se në të njëjtën kohë glukagoni stimulon fosforilimin dhe inaktivizimin e fosfotazës së reduktazës HMG-CoA, duke e mbajtur kështu reduktazën HMG-CoA në një gjendje joaktive fosforiluese. Si rezultat, sinteza e kolesterolit në periudhën pasabsorbimit dhe gjatë agjërimit frenohet nga sinteza endogjene. Nëse përmbajtja e kolesterolit në ushqim u soll në 2%, atëherë sinteza e kolesterolit endogjen u ul ndjeshëm. Por ndërprerja e plotë e sintezës së kolesterolit nuk ndodh. Shkalla e frenimit të biosintezës së kolesterolit nën ndikimin e kolesterolit që vjen nga ushqimi ndryshon nga personi në person. Kjo tregon individualitetin e proceseve të formimit të kolesterolit. Duke zvogëluar intensitetin e sintezës së kolesterolit, është e mundur të zvogëlohet përqendrimi i tij në gjak. Nëse ekuilibri midis konsumit të kolesterolit me ushqim dhe sintezës së tij në trup nga njëra anë dhe sekretimit të acideve biliare dhe kolesterolit nga ana tjetër është i prishur, përqendrimi i kolesterolit në indet dhe gjaku ndryshon. Pasojat më të rënda shoqërohen me një rritje të përqendrimit të kolesterolit në gjak (hiperkolesterolemia), ndërsa gjasat e zhvillimit të aterosklerozës dhe kolelithiasis rritet. Hiperkolesterolemia familjare (HCS) - kjo formë është shumë më e zakonshme - rreth 1 pacient për 200 persona. Një defekt i trashëguar në HCS është një shkelje e thithjes së LDL nga qelizat, dhe për këtë arsye, një rënie në shkallën e katabolizmit LDL. Si rezultat, përqendrimi i LDL në gjak rritet, si dhe kolesterolit pasi ka shumë në LDL. Prandaj, me HCS, depozitimi i kolesterolit në inde, në veçanti në lëkurë (xanthoma), në muret e arterieve është karakteristik. Pengimi i sintezës së reduktazës HMG-CoA Produkti përfundimtar i rrugës metabolike të kolesterolit. Ajo zvogëlon shkallën e transkriptimit të gjenit HMG-CoA reductase, duke penguar kështu sintezën e vet. Mëlçia po sintetizon në mënyrë aktive acidet biliare nga kolesteroli, dhe për këtë arsye acidet biliare frenojnë aktivitetin e gjenit HMG-CoA reductase. Meqenëse reduktaza HMG-CoA ekziston pas sintezës së rreth 3, frenimi i sintezës së këtij enzima të kolesterolit është një rregullim efektiv.
|